Home
Shop
Services
News
Success Stories
About Us
Courses
Contact us
Category/Dna
Atto
Biocoupler Plant Propagation
Suppliers
PSEP Assay
2012/12/14/peptide-based-vaccines-for-immunotherapies
products
category/comparative
2014/06
products/oligos
IVD
0
+1 (650) 555-0111
Sign in
Contact Us
0
Home
Shop
Services
News
Success Stories
About Us
Courses
Contact us
Category/Dna
Atto
Biocoupler Plant Propagation
Suppliers
PSEP Assay
2012/12/14/peptide-based-vaccines-for-immunotherapies
products
category/comparative
2014/06
products/oligos
IVD
+1 (650) 555-0111
Sign in
Contact Us
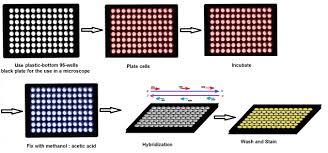
KD Linse — By Klaus D. Linse Fluorescence In Situ Hybridization (FISH) for Telomere Length Quantification or Q-FISH evolves into High-Throughput Q-FISH.